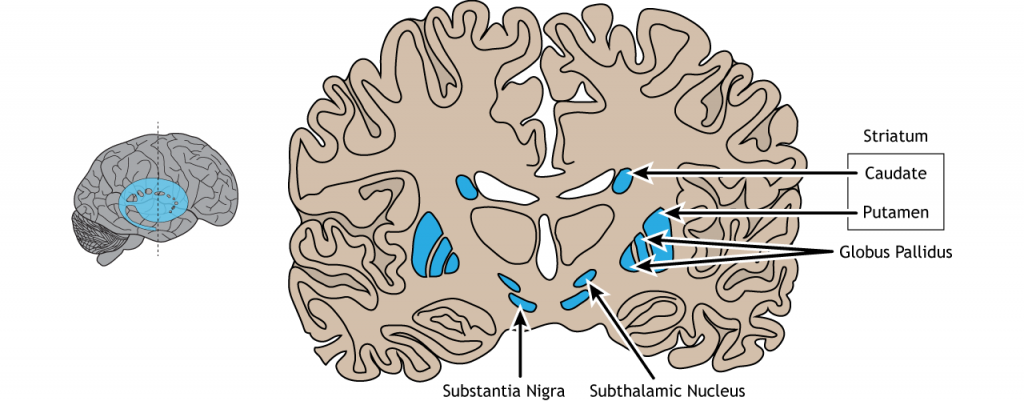
The basal ganglia are a group of subcortical nuclei, meaning groups of neurons that lie below the cerebral cortex. The basal ganglia is comprised of the striatum, which consists of the caudate nucleus and the putamen, the globus pallidus, the subthalamic nucleus, and the substantia nigra The basal ganglia are primarily associated with motor control, since motor disorders, such as Parkinson’s or Huntington’s diseases stem from dysfunction of neurons within the basal ganglia. For voluntary motor behavior, the basal ganglia are involved in the initiation or suppression of behavior and can regulate movement through modulating activity in the thalamus and cortex. In addition to motor control, the basal ganglia also communicate with non-motor regions of the cerebral cortex and play a role in other behaviors such as emotional and cognitive processing.
The majority of information processed by the basal ganglia enters through the striatum. The principal source of input to the basal ganglia is from the cerebral cortex. This input is glumatergic and therefore, excitatory. The substantia nigra is also a region with critical projections to the striatum and is the main source of dopaminergic input. Dopamine plays an important role in basal ganglia function. Parkinson’s disease results when dopamine neurons in the substantia nigra degenerate and no longer send appropriate inputs to the striatum. Dopamine projections can have either excitatory or inhibitory effects in the striatum, depending on the type of metabotropic dopamine receptor the striatal neuron expresses. Dopamine action at a neuron that expresses the D1 receptor is excitatory. Dopamine action at a neuron that expresses the D2 receptor is inhibitory.
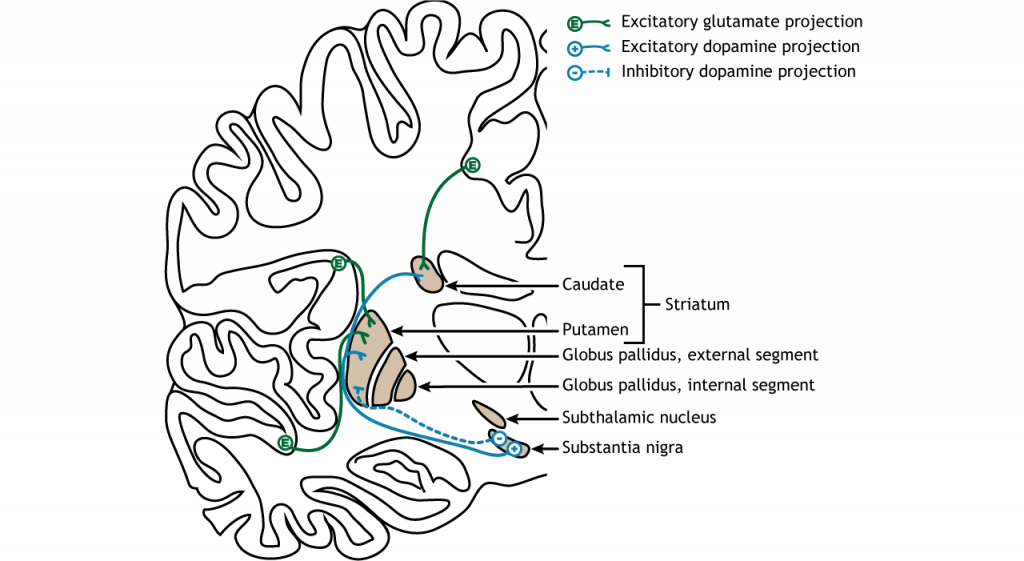
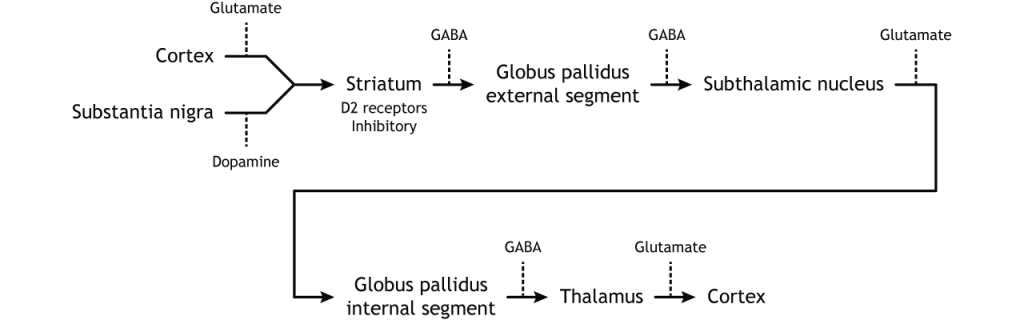
The primary output region of the basal ganglia is the internal segment of the globus pallidus. This region sends inhibitory GABAergic projections to nuclei in the thalamus. This inhibitory output has a tonic, constant firing rate, which allows the basal ganglia output to both increase and decrease depending on the situation. The thalamus then projects back out to the cerebral cortex, primarily to motor areas.
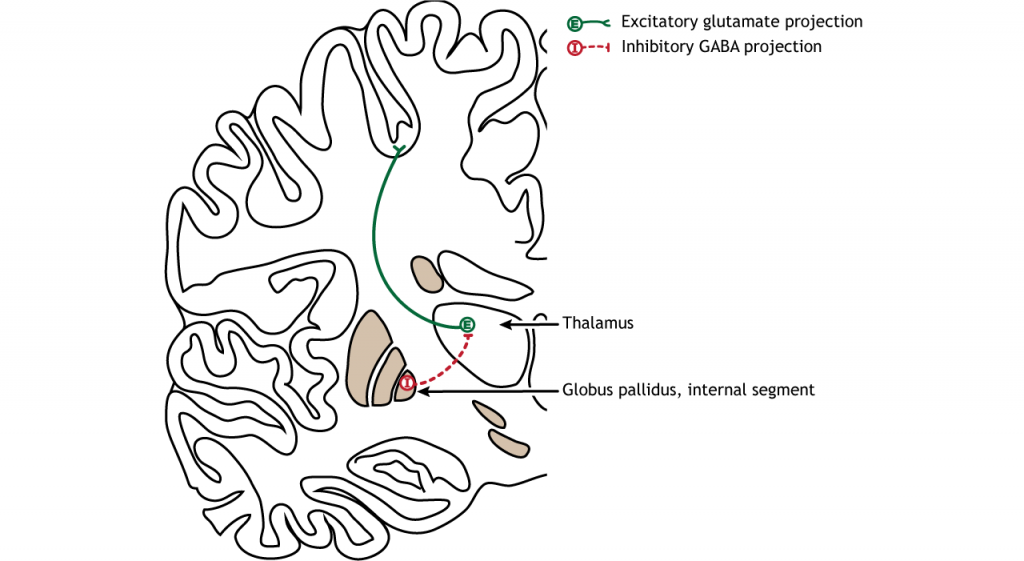

There are multiple connections within the basal ganglia structures as well. For motor control, there are two main circuits: the direct pathway and the indirect pathway. These circuits have opposing actions when activated by cortical neurons. The circuits are also modulated by dopamine release by the substantia nigra into the striatum. It is believed that the different control mechanisms allow a finely tuned balance between the direct and indirect circuits, which allows for refined control of movement.
The direct pathway begins in the striatum, which sends inhibitory projections to the internal segment of the globus pallidus (GPi). The GPi then sends inhibitory output to the thalamus.
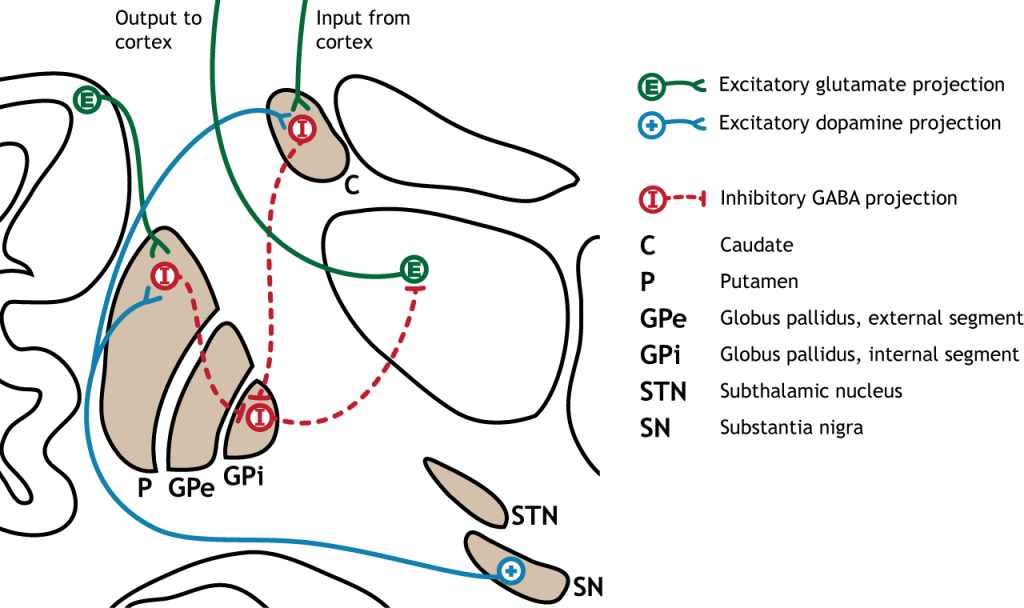
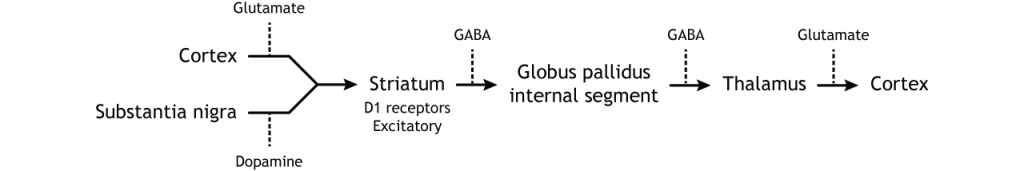
When input from either the cortex or substantia nigra increases in intensity, the direct pathway is activated. The neurons in the striatum involved in the direct pathway express the D1 metabotropic dopamine receptor, and the activation of this receptor is excitatory. Therefore, projections from both the cortex and the substantia nigra activate the neurons in the striatum. Those neurons are inhibitory and release GABA onto the internal segment of the globus pallidus (GPi). As described above, the neurons in the GPi are inhibitory, releasing GABA onto the thalamus. Activation of the striatum neurons inhibit the neurons in the GPi, releasing the inhibition on the thalamus. Inhibition of an inhibitory region is called disinhibition. Therefore, the activation of the direct pathway results in increased output from the thalamus because it is disinhibited.
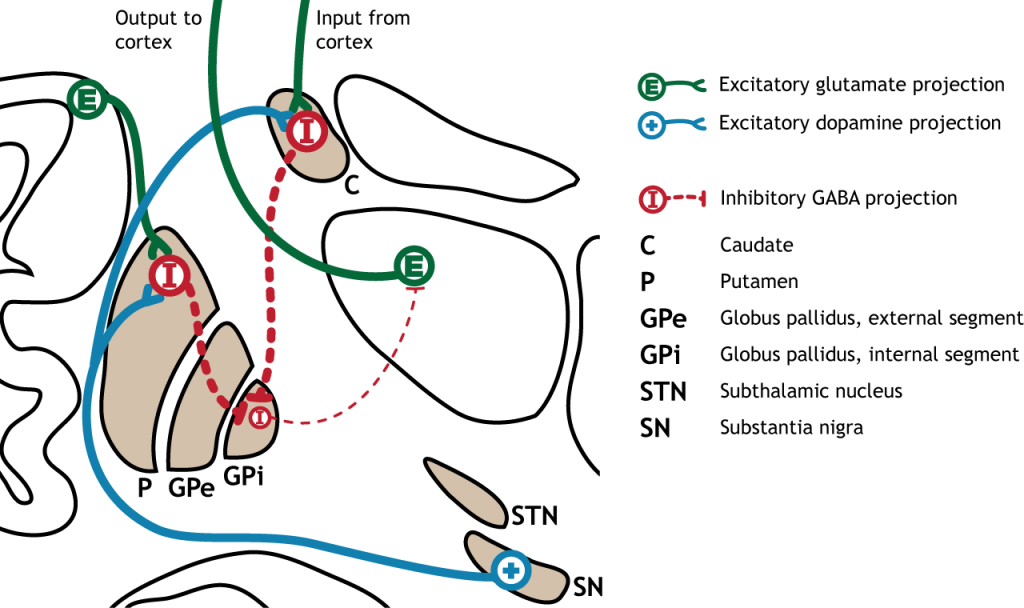
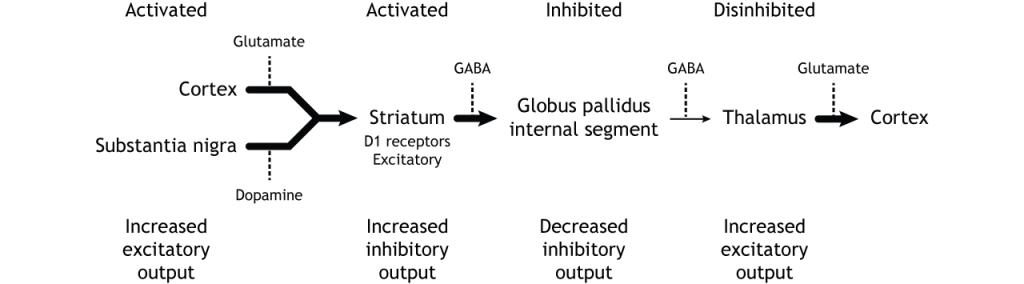
The indirect pathway is a little more complex. Like the direct pathway, input into the basal ganglia arises from the cortex and substantia nigra, but there are more internal connections within the basal ganglia that what occurs in the direct pathway. Inhibitory neurons in the striatum involved in the indirect pathway project to the external segment of the globus pallidus (GPe). GABA-ergic neurons in the GPe project to the subthalamic nucleus, which then sends excitatory output to the GPi, which outputs to the thalamus.
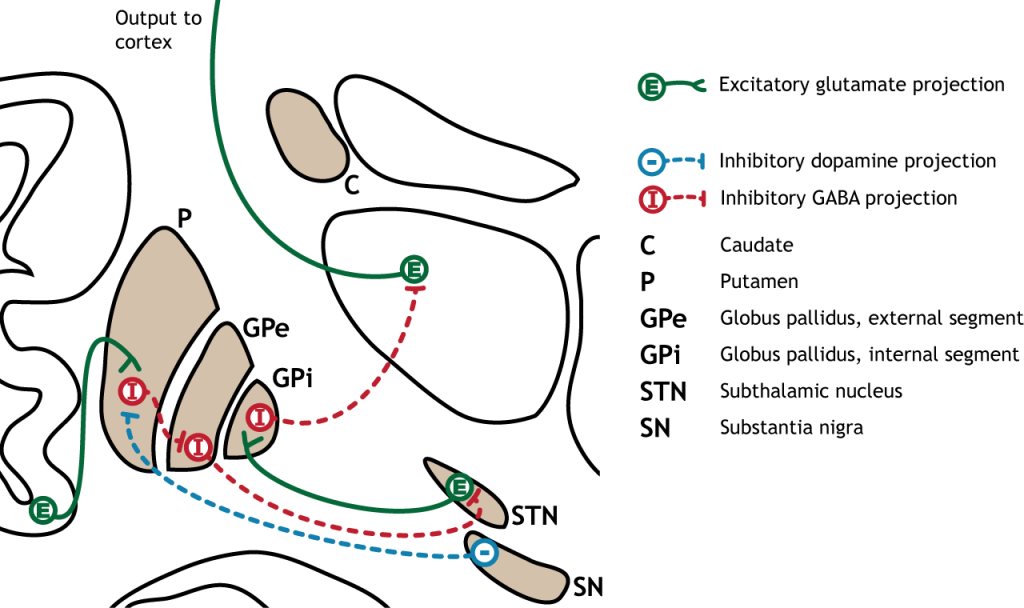
The indirect pathway is activated by excitatory cortical input, activating the inhibitory striatal neurons. This leads to inhibition of the GPe neurons, resulting in disinhibition of the excitatory neurons in the subthalamic nucleus. The excitatory output from the subthalamic nucleus to the GPi increases inhibition of the thalamus, leading to decreased thalamic output to the cortex.
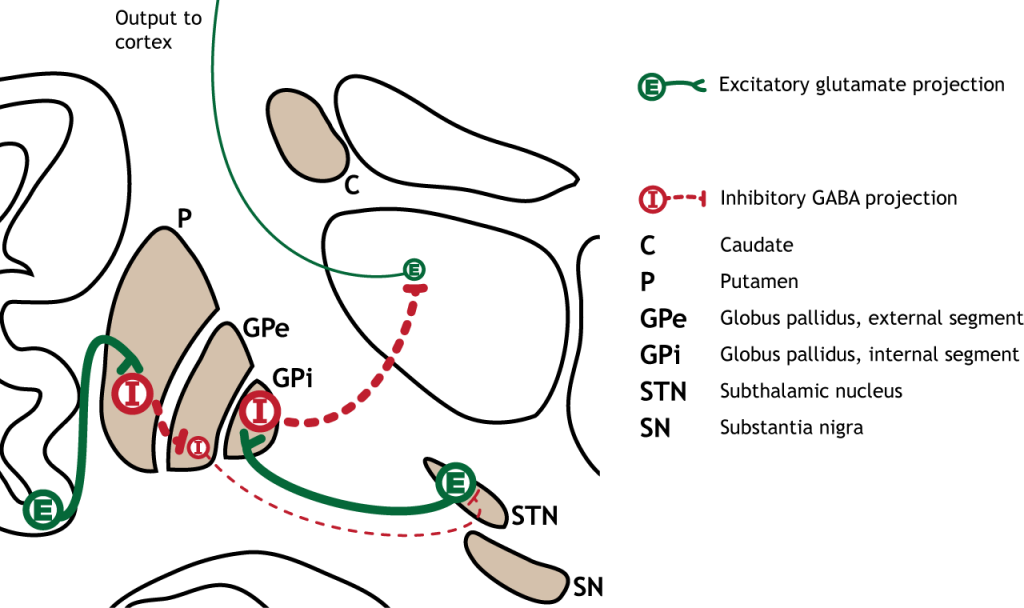
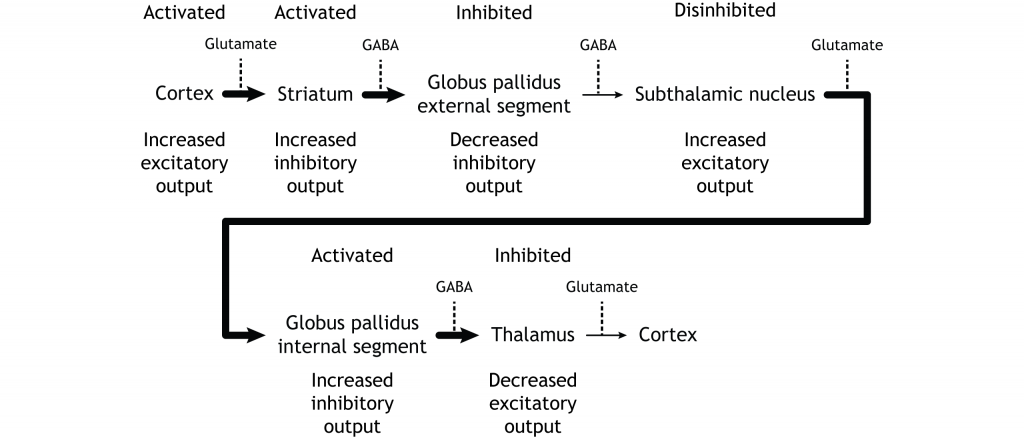
The indirect pathway can be inhibited by dopamine release from the substantia nigra. The neurons in the striatum involved in the indirect pathway express the D2 metabotropic dopamine receptor. The activation of this receptor is inhibitory. If the indirect pathway is inhibited by dopamine projections from the substantia nigra, the inhibitory striatal neurons are inhibited. This leads to disinhibiton of the GPe neurons, resulting in inhibition of the excitatory neurons in the subthalamic nucleus. This decreased excitatory output to the GPi decreases inhibition of the thalamus, leading to increased thalamic output to the cortex.
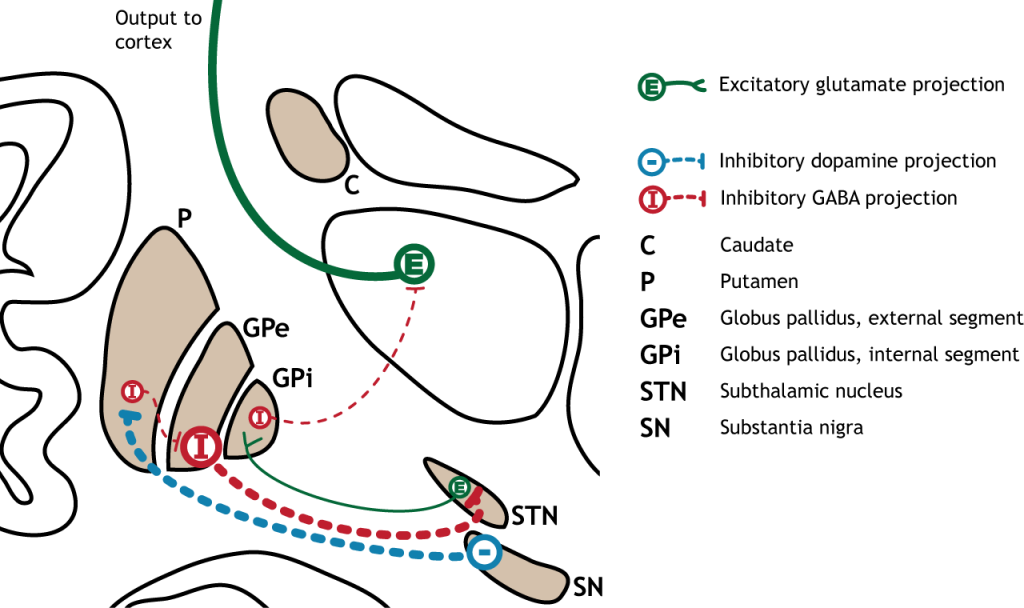
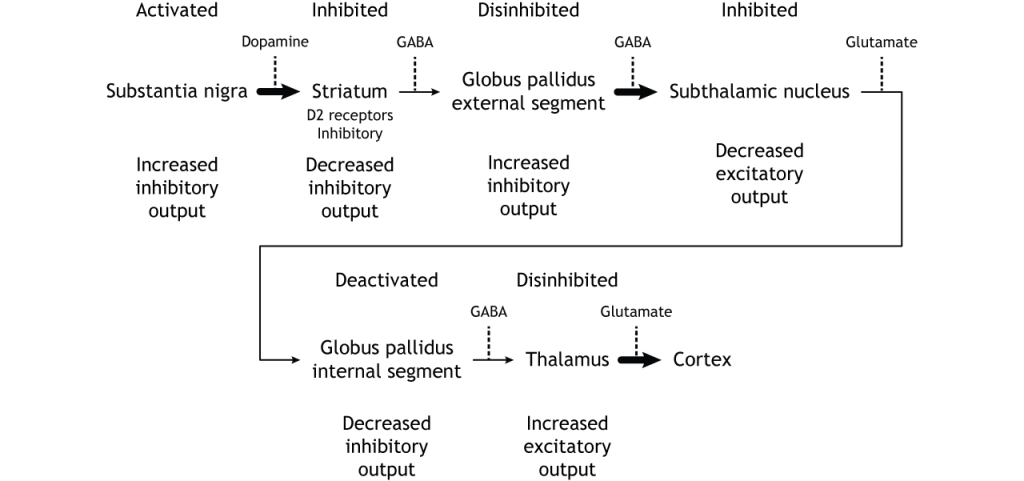
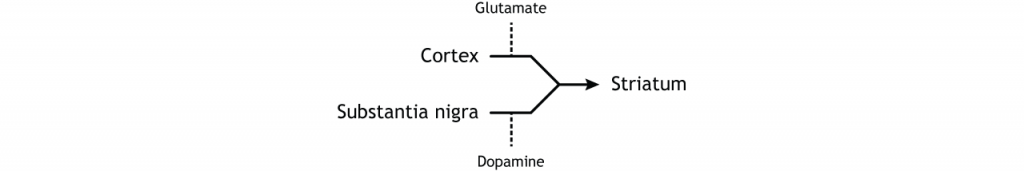
To put it all together, there is input to the striatum from two different locations: cortex (glutamate) and substantia nigra (dopamine).
It is the combination of these pathway that allows for precise control of motor movement.
There are multiple circuits that pass through the basal ganglia:
Although the circuits each use different circuits within the basal ganglia, the general loop is the same: cortical input to the striatum leads to internal processing within the basal ganglia structures. Basal ganglia output projects from the pallidum to the thalamus, which then projects back to the cortex. It is important to recognize that the basal ganglia plays an important role in a number of functions. For example, medications that are used to treat Parkinson’s can sometimes lead to the presentation of impulse control disorders, a result of dopaminergic changes in the limbic loop through the basal ganglia.
![]()